WSRC-MS-2003-00337
Using Sulfate-Amended Sediment Slurry
Batch Reactors to Evaluate Mercury Methylation
Sarah M. Harmon and G.Thomas Chandler
University of South Carolina
Arnold School of Public Health
Department of Environmental Health Sciences
Columbia, South Carolina 29208, USA
Jeffrey K. King
Applied Technology and Management, Inc.
Savannah, GA 31405
John B. Gladden
Westinghouse Savannah River Company
Aiken, South Carolina, 29808
Lee A. Newman
University of Georgia
Savannah River Ecology Laboratory
Drawer E
Aiken, SC 29808
Abstract
In the methylated form, mercury represents a concern to public health primarily through the consumption of contaminated fish tissue. Research conducted on the methylation of mercury strongly suggests the process is microbial in nature and facilitated principally by sulfate-reducing bacteria. This study addressed the potential for mercury methylation by varying sulfate treatments and wetland-based soil in microbial slurry reactors with available inorganic mercury. Under anoxic laboratory conditions conducive to growth of naturally occurring sulfate-reducing bacteria in the soil, it was possible to evaluate how various sulfate additions influenced the methylation of inorganic mercury added to overlying water. Treatments included sulfate amendments ranging from 25 to 500 mg/L (0.26 to 5.2 mM) above the soil's natural sulfate level. This study also provided an assessment of mercury methylation relative to sulfate-reducing bacterial population growth and subsequent sulfide production. Mercury methylation in sulfate treatments did not exceed that of the non-amended control during a 35-day incubation. However, increases in methylmercury concentration were linked to bacterial growth and sulfate reduction. A time lag in methylation in the highest treatment correlated with an equivalent lag in bacterial growth.
Keywords: Mercury, Methylmercury, Sulfate-Reducing Bacteria, Sulfate, Sulfide
Introduction
Mercury is a common environmental contaminant which becomes much more toxic in its methylated form because of methylmercury's ability to easily cross cell membranes, accumulate in biological tissues, then biomagnify through the food chain [1,2]. From the standpoint of human health risk, the accumulation of methylmercury in edible fish tissue has resulted in fishing restrictions, numerous health advisories, and much public apprehension [3-5].
In wetland environments that receive anthropogenic mercury contributions, the activity of sulfate reducing bacteria is especially important, and the effect of sulfate-reducing bacterial metabolism is two-fold:
- Methylmercury forms if sulfate reducers respire in the presence of a bioavailable species of inorganic mercury [6-10], as mercury methylation occurs through a side reaction within the bacteria's normal metabolic pathway [11-13].
- Sulfide is produced via bacterial respiration and the subsequent reduction of sulfate. In turn, the bioavailability of inorganic mercury to sulfate reducers is largely controlled by porewater sulfide concentration [14-16].
Historically, direct precipitation as solid HgS as well as adsorption to solid phases were thought to control dissolved mercury concentration. While the reaction of sulfide with Hg2+ to produce insoluble cinnabar (HgS) certainly decreases mercury availability for methylation [17], there is believed to be a range of low sulfide concentrations which may actually enhance mercury bioavailability. Several studies have noted that the concentration of dissolved (bioavailable) inorganic mercury in sulfidic porewaters increases along with sulfide concentration to a certain degree [18,19]. Benoit et al. [14,15,18] hypothesized that, under low sulfidic conditions, the availability of mercury for methylation is increased by the formation of a neutral dissolved mercury-sulfide complex (HgS0). The concept behind the HgS0 hypothesis is that this uncharged chemical species readily enters the bacterial cells through passive diffusion. Once inside, cellular metabolism will result in methylation of the mercury ion. As the concentration of sulfide increases, an equilibrium shift will result in a decrease in HgS0 as the neutral species is replaced with charged disulfide complexes [14,15]. Mercury methylation in a pure culture experiment has been shown to decrease about fourfold as sulfide concentration increased from 10-6 to 10-3 M [16].
So while sulfate-reducing bacteria are respiring and producing sulfide that will sequester available metal cations (including inorganic mercury) into a non-toxic precipitate, they can also methylate a portion of this same mercury pool into a more toxic bioavailable form. Mercury methylation has been linked to the activity of sediment sulfate-reducing bacteria in the Florida Everglades [17], the Quabbin Reservoir in Massachusetts [10], Pallette Lake in northern Wisconsin [20], and the Patuxent River Estuary in Maryland [18].
Research in the area of constructed wetland technologies has attempted to capitalize on the formation of sulfide in order to enhance the removal of metal cations from industrial outfalls, discharges, and other surface water input [21]. Under normal freshwater conditions, sulfate concentrations are relatively low (5x10-5 to 4.5x10-4 M) [22], and sulfate-reducing bacteria would not be expected to out compete other microbial organisms. In such systems, mercury methylation mediated by sulfate reducers would be at a minimum. Treatment systems developed for the enhanced production of sulfide, however, typically have soil sulfate amendments (e.g., gypsum) which change the natural underlying sediment chemistry. On a microbial level, the augmentation of soils with sulfate potentially favors selection of sulfate-reducing bacteria resulting in population growth and an overall increase in bacterial sulfate reduction. Subsequently, increased sulfate reduction potentially leads to an increase in methylmercury production, provided mercury is bioavailable [23]. Significant efforts must be made to ensure that constructed wetlands systems augmented with sulfur-derived compounds are not increasing the concentration of a more harmful mercury species once treatment is presumed complete. This study evaluates the dynamics of the sulfate-mercury relationship in a representative soil to fully assess the risk vs. benefit of these amendments.
Various sulfate treatments and soil from a proposed wetland treatment area were combined with biologically available mercury in sediment slurry reactor systems, which provided a method for observing the activity of indigenous sulfate-reducing bacteria under simulated anaerobic wetland conditions. Slurry batch reactor systems were constructed using an aqueous media that was conducive to the growth of sulfate-reducing bacteria. These systems were placed under anoxic laboratory conditions and examined to learn the relationship between soil treatment, potential methylation of inorganic mercury, and sequestration of metals from the overlying water. Additionally, this study provides insight into mercury methylation with respect to the population growth of sulfate-reducing bacteria and subsequent sulfide production.
Materials and Methods
The laboratory study was designed to simulate anaerobic wetland sediment conditions through the use of N2/CO2-purged batch reactors incubated in an anaerobic chamber. Reactors containing a sediment slurry (simulated hydrosoil, water, and a series of sulfate treatments) were spiked with aqueous mercury. The experiment included four sulfate treatments with three replicates per treatment. Aliquots were periodically removed and measured for sulfate, sulfide, total mercury, methylmercury, and sulfate-reducing bacterial population estimates.
A 35-day experimental duration was selected based upon preliminary data and sampling limitations within the experimental microcosms. The volume of sediment slurry reactors was limited and, therefore, so was the number of samples which could be taken without compromising the experiment. In preliminary unpublished experiments, 35 days was the duration required for bacterial populations to deplete all of the free sulfate present in the slurry vessels. It was assumed that the influence of sulfate additions to the sediment slurries could be fully characterized within this time period.
Sediment Slurry Reactor Systems
Slurry reactor vessels were constructed using 300 mL Fleakerâ Beakers (Corning) with gas-tight caps fitted with three glass-tube ports through the cap (Figure 1). One stop-cocked tube allowed addition of N2/CO2 (9:1) through the slurry during sample preparation to achieve initial anoxic conditions within the reactor vessel. A second tube with a check valve served as a gas release port, and the remaining tube served as a sampling port.
Reactors contained simulated hydrosoil suspended in a slurry of aqueous media with varying sulfate concentration. The media was modified from [24] to control the sulfate concentration while still providing nutrients for the support of bacterial consortia which would be expected in a healthy wetland with actively growing vegetation. Media components were mixed into ultrapure water as follows: lactic acid (5 mL/L), CaCl2· 2H2O (60 mg/L), MgCl2· 6H2O (400 mg/L), sodium citrate (300 mg/L), K2HPO4 (640 mg/L), NH4Cl (1000 mg/L). The pH was adjusted to 6.8 using 4N NaOH. Once mixed, sulfate was added in the form of CaSO4· ½H2O to achieve nominal concentrations of 25, 100, and 500 mg/L. The control had no additional sulfate added; however, once mixed with the hydrosoil, some sulfate was present due to that which was naturally available in the soil. Actual sulfate concentrations were determined via ion chromatography. Media was purged with the N2/CO2 gas for 30 min prior to sediment addition.
The simulated hydrosoil was made by mixing a sample of the soil proposed for use in wetland construction with an organic component to simulate conditions in the constructed wetland after approximately one year of operation. The organic component consisted of a contaminant-free sediment retrieved from Flamingo Bay on SRS and partially decomposed plant material (Schoenoplectus sp.). Percent moisture was determined for both the soil and the organic component, and dry weight was calculated. Hydrosoil was then mixed to achieve a 1:3 soil to organic component ratio based on this dry weight estimate. Hydrosoil was homogenized then divided among reactors at a rate of 50 g per beaker. Wet weight of soil in each reactor was recorded. An aliquot of the hydrosoil mixture was set aside for dry weight measurement. This data was used to estimate the dry weight content of soil in each reactor and for the dry weight normalization of sediment mercury data. The deaerated media with sulfate addition was then added at a rate of 200 mLs per beaker. The reactors were thoroughly mixed then transferred into an anaerobic chamber. Each reactor was mixed twice per day and maintained at 27° C throughout the incubation.
Two of the replicates were sampled five times over the incubation period for mercury, and methylmercury. A third replicate was used to monitor sulfate, sulfide, and bacterial populations on more frequent basis. The use of third replicate allowed for frequent sampling of these parameters without compromising volume in the mercury-sampled reactors. Mercury concentrations from this third replicate were not considered in the final data analysis.
Dissolved inorganic mercury (Hg2+) was added as mercuric nitrate in 11 stages over the 35-day incubation period to reach a final cumulative concentration of 1 m g Hg/g(dry wt). This concentration was selected to mirror actual field conditions observed in an earlier study [25] and are believed to be consistent with the expected constructed wetland conditions. Each mercury addition was 3 m g Hg per test vessel for a total of 33 m g by the end of the incubation period. Mercury was added in stages so that bioavailable inorganic mercury would be present for bacterial populations over most of the period when bacterial activity was judged to be highest. Mercury was added on Days 9, 13, 15, 17, 20, 21, 22, 23, 24, 27, and 29.
Sediment slurries were sampled for mercury and methylmercury by pulling 30-mL of the slurry out of the test vessel via syringe then transferring into polypropylene centrifuge tubes. Samples were briefly centrifuged before the overlying water was poured off for separate analysis. The sediment pellet was immediately frozen at -20°C.
Chemical Analyses
Total And Methylmercury in Sediment and Water
Low-level methylmercury analysis in water and sediment samples was preformed using a technique modified from US EPA Methods 1630 [26]. Humics were removed from the samples through subboiling distillation of acidified samples [27]. Mercury in the distillate was then ethylated with sodium tetraethylborate and purged with argon onto a trap packed with Tenax [28]. The Tenax was flash heated in a stream of argon to release the mercury which was speciated chromatographically [29], combusted to Hg0, and measured using cold vapor atomic fluorescence spectrometry (CVAFS)[30]. The CVAFS mercury analyzer was a Tekron Model 2500 (Tekron Inc, Ontario, Canada) and the integrator was a Hewlett-Packard Model HP3394A (Hewlett Packard Co., Boise, ID). The detection limit was 0.02 ng/L as Hg.
Low-level total mercury analysis in water samples followed US EPA Method 1631 [31]. For this method, all species of mercury in the sample were oxidized to Hg2+ through the addition of BrCl, then reduced to volatile Hg0 by reacting the sample with SnCl2. The Hg0 was separated from solution by purging with argon onto a gold-coated sand trap [32]. The trapped mercury was then thermally desorbed from the gold trap into an argon stream that carried released Hg0 into the cell of the CVAFS mentioned above for quantification [30].
Low-level total mercury in sediment samples was measured via Inductively-Coupled Plasma Mass Spectrometry (ICP-MS) using the cold-vapor reduction, isotope dilution method described by [33]. Sediment slurry samples (0.5 g) were acidified with nitric acid, spiked with 0.5 mL of an enriched 201Hg isotope solution, then microwave digested. Aliquots of the digested samples were then reacted with a combined solution of potassium hydroxide (0.1%) and sodium borohydride (5%) for the reduction and volatilization of all mercury species which were carried into the instrument in an argon stream. Total mercury concentration in the sample was determined based on the ratio of 202Hg/201Hg. This analysis was carried out using a VG Plasma Quad 3 ICP-MS (Thermo VG Scientific, West Sussex, England).
Other Analyses
Samples for sulfate analysis were filtered through 0.45 m m AcrodiscÒ syringe filters (Gelman Laboratory, Ann Arbor, MI) then measured using a Dionex model DX-120 ion chromatograph with an IonPac As14 separation column (Dionex Corp, Sunnyvale, CA). Sulfide samples were diluted 1:1 with a sulfide anti-oxidant buffer [34] which served as a preservative when the sample was exposed to air. Total sulfide concentrations were determined via silver-sulfide ion-specific electrode and meter (Thermo Orion, Beverly, MA). A Hach One pH meter was used to determine pH (Hach, Loveland, CO.).
Bacterial Enumeration
A most probable numbers (MPN) technique was used to estimate the number of viable sulfate reducing bacteria in each reactor set over time. This procedure was done with a series of six to eight descending 10-fold dilutions with five replicates per dilution. Enrichment media was prepared following standard protocol [35], and testing was performed in Hungate anaerobic culture tubes (Bellco Glass, Inc., Vineland, NJ). Culture tubes were then inoculated from the experimental reactors using aseptic technique in an anaerobic chamber. After inoculation, tubes were stored for one month before determining final scores. Tubes were scored positive by the production of a black FeS precipitate, indicating the presence of active sulfate-reducing bacteria. Standard MPN evaluation tables and 95% confidence intervals [35] were used to estimate the number of viable sulfate reducing bacteria in each reactor set. Both positive and negative controls were included to ensure viability of the culture media and proper inoculation technique.
Results and Discussion
Over the 35-day time period of this experiment, sediment methylmercury concentrations increased in all sulfate treatments (Figure 2). However, none of the sulfate treatments produced methylmercury in concentrations significantly greater than that of the non-sulfate-amended control. This trend was evident in both the sediment slurry (Figure 2) and in the overlying water (Figure 3). Sediment methylmercury concentrations in the highest sulfate treatment were notably lower on Days 30 and 35, and this trend was also noted in the overlying water on the final day of the experiment. This data would lead to the expected conclusion that a higher initial sulfate concentration (≥500 mg/L) would result in lower mercury methylation rates; however, further examination of the sulfate chemistry and bacterial dynamics give greater insight into the mechanisms contributing to this result.
When mercury methylation is considered in conjunction with bacterial population dynamics and sulfate chemistry (Figure 4 A-D) it is evident that mercury methylation in these reactor systems is related to active sulfate-reducing bacteria growth and subsequent reduction of sulfate to sulfide. It should be noted that while the control treatment was not amended with additional sulfate, there was enough sulfate already present in the soil to support sulfate reducers. In the control and two lowest sulfate treatments (Figure 4 A-C), inorganic mercury was added to actively growing bacterial populations, and methylation occurred between the initial mercury spike on day 9 and the next sampling period on day 21. In the highest sulfate treatment (Figure 4D), bacterial growth was delayed, as was measurable mercury methylation. The reason for delay in bacterial growth in the highest sulfate treatment is unclear, but was likely due to chemical conditions in the media. As stated earlier, sulfate-reducing bacteria have long been implicated as the primary methylators of mercury under anaerobic conditions. A recent study using pure cultures of Desufobulbus propionicus (1pr) have shown that sulfate-reducing bacteria cells must be present before methylation will occur and that methylation rates were highest when inorganic mercury was spiked into actively growing cultures [16]. Furthermore, a kinetic relationship between sulfate reduction and mercury methylation rate [23,36] provides further evidence of a pairing of these two processes.
Since these slurry reactors are closed systems which are both carbon and sulfate limited, sulfate-reducing bacteria population growth would be expected to plateau and then decline as resources are depleted. This trend can be noted in the all sulfate treatments (Figures 4 A-D). Even though sulfate-reducing bacteria populations were no longer in log-phase growth at the experiments termination, methylmercury continued to increase as the methylation was sustained by active cells. It is possible that inorganic mercury was more bioavailable due to the sulfide production. Moreover, pure culture studies have shown that Desufobulbus propionicus (1pr) will methylate HgS0 under fermentative conditions [16,37], and this could also explain the continued methylation observed in the first three sulfate treatments after sulfate was depleted.
Mercury methylation in the non-amended control appears to level off between days 30 and 35 (Figure 4A). This may be attributed to both a decline in bacterial population and bacterial demethylation. As with the mercury methylation process, demethylation has also been linked to the activity of sulfate-reducing bacteria [38] and has been demonstrated in pure cultures of Desulfovibrio desulfuricans LS [39].
Data from this experiment also supports the idea that a low-concentration sulfide pool may make mercury more bioavailable as a neutral HgS0 species for uptake by the bacteria. In each sulfate treatment, formation of sulfide (10-7M to 10-4M) initiated the period of mercury methylation (Figure 5 A-D). In the highest sulfate treatment (Figure 5D) where bacterial growth was delayed for 15 days, methylation was not observed for an additional 15 days, and this was only after sulfide reached the maximum concentration measured in this experiment (3.63x10-4 M).
As with all microcosm studies, these slurry reactor systems cannot fully mimic in situ wetland conditions due to size restrictions, nutrient limitations, and the lack of natural variability present in actual wetland environments. They did, however, provide a controlled glimpse into the dynamics of the bacterial consortia present in this particular soil and offered a brief evaluation of the related sulfur and mercury biogeochemistry. Sulfate amendments ranging from 25 to 500 mg/L (0.26 to 5.2 mM) above the soil's natural sulfate level did not increase the mercury methylation over that of the control during this 35-day experiment. While mercury methylation was observed in all treatments, there was a lag time in the highest treatment that was likely linked to an equivalent lag in bacterial growth. While it is difficult to discern a recommendation based solely on this data, this experiment supplies a starting point for the evaluation of the risk vs. benefit of sulfur additions to this particular wetland treatment system. Field studies will add to the conclusions presented here by providing conditions where bacterial mediated mercury methylation and demethylation can be evaluated in conjunction with a constantly changing wetland environment.
Acknowledgement
The authors would like to thank Debbie Wells, James Bowers, Robert Ray, Morris Jones, Diane Addis, Pam Mckinsey, and Chuck Turick for analytical and technical assistance. All mercury samples were analyzed at the Skidaway Institute of Oceanography, Savannah, GA. This work was funded by the U.S. Department of Energy and Westinghouse Savannah River Company through a research program administered by the Oak Ridge Institute for Science and Education.
References
- Kelly CA, Rudd JWM, St. Louis VL, Heyes A. 1995. Is total mercury
concentration a good predictor of methyl mercury concentration in aquatic
systems. Water Air Soil Pollut 80: 715-724.
- Bodaly RA, St. Louis VL, Paterson MJ, Fudge RJP, Hall BD, Rosenberg
DM, Rudd JWM. 1997. Bioaccumulation of mercury in the aquatic food chain
in newly flooded areas. In Sigel A, Sigel H, eds, Metal Ions in Biological
Systems, Volume 34: Mercury and Its Effects on Environment and Biology,
Marcel Dekker Inc, New York, NY. pp 259-287.
- NRC. 2000. Toxicological Effects of Methylmercury. National
Research Council. National Academy Press. Washington, D.C.
- US EPA. 1997. Mercury Report to Congress. Office of Air Quality and
Standards. Washington, DC.
- US EPA. 2002. Update: National Listing of Fish and Wildlife Advisories.
US EPA Fact Sheet. EPA-823-F-02-007. Office of Water, Washington, DC.
May 2002. http://www.epa.gov/waterscience/fish/advisories/factsheet.pdf
- Compeau G, Bartha R. 1985. Sulfate reducing bacteria: Principal methylators
of mercury in anoxic estuarine sediments. Appl Environ Microbiol
50:498-502.
- Compeau G, Bartha R. 1987. Effect of salinity on mercury methylating
activity of sulfate reducing bacteria in estuarine sediments. Appl
Environ Microbiol 3:261-265.
- Devereux R, Winfrey MR, Winfrey J, Stahl DA. 1996. Depth profile of
sulfate-reducing bacterial ribosomal RNA and Hg methylation in an estuarine
sediment. FEMS Microbiol Ecol 20:23-31.
- Gilmour CG, Henry EA. 1991. Mercury methylation in aquatic systems
affected by acid deposition. Environ Pollut
71:131-169.
- Gilmour CG, Henry EA, Mitchell R. 1992. Sulfate stimulation of mercury
methylation in freshwater sediments. Environ Sci Techno 26:2281-2287.
- Berman M, Chase T, Jr., Bartha R. 1990. Carbon flow in mercury biomethylation
by Desulfovibrio desulfuricans. Appl Environ Microbiol
56:298-300.
- Choi SC., Bartha R. 1993. Cobalamin-mediated mercury methylation by
Desulfovibrio desulfuicans. Appl Environ Microbiol 59:290-295.
- Choi SC, Chase T, Jr., Bartha T. 1994. Metabolic pathways leading
to mercury methylation in Desulfovibrio desulfuicans LS. Appl
Environ Microbiol 60:4072-4077.
- Benoit JM, Gilmour CC, Mason RP, Heyes A. 1999a. Sulfide Controls
on Mercury Speciation and Bioavailability to Methylating Bacteria in
Sediment Pore Waters Environ Sci Technol 33:951-957.
- Benoit JM, Mason RP, Gilmour CC. 1999b. Estimation of mercury-sulfide
speciation in sediment pore waters using octanol water partitioning
and implications for availability to methylating bacteria. Environ
Toxicol Chem 18:2138-2141.
- Benoit JM, Gilmour CC, Mason RP. 2001a. Aspects of bioavailability
of mercury for methylation in pure cultures of Desulfobulbus propionicus
(1pr3). Appl Environ Microbiol 67:51-58.
- Gilmour CG, Riedel GS, Ederington MC, Bell JT, Benoit JM, Gill GA,
Stordal MC. 1998. Methylmercury concentration and production rates across
a trophic gradient in the northern Everglades. Biogeochemistry
40:327-345.
- Benoit JM, Gilmour CC, Mason RP, Riedel GS, Riedel GF. 1998.
Behavior of mercury in the Patuxent River Estuary. Biogeochemistry
40:249-265.
- Bloom NS, Gill GA, Cappellino S, Dobbs C, McShea L, Driscoll C, Mason
R, Rudd J. 1999. Speciation and cycling of mercury in Lavaca Bay, Texas
sediments. Environ Sci Technol 33:7-13.
- Watras CJ, Bloom NS, Claas SA, Morrison KA, Gilmour CC, Craig SR.
1995. Methylmercury production in the anoxic hypolimnion of a dimictic
seepage lake. Water Air Soil Pollut 80:735-745.
- Dvorak DH, Hedin RS, Edenborn HM, McIntire PE. 1992. Treatment of
metal contaminated water using bacterial sulfate reduction: results
from pilot-scale reactors. Biotechnology and Bioengineering 40:609-616.
- Feng J, Hsieh YP. 1998. Sulfate reduction in freshwater wetland soils
and the effects of sulfate and substrate loading. J Environ Qual
27:968-972.
- King JK, Saunders FM, Lee RF, Jahnke RA. 1999. Coupling mercury methylation
rates to sulfate reduction rates in marine sediments. Environ Toxicol
Chem 18(7):1362-1369.
- Widdel F, Bak F. 1992. Gram-Negative Mesophilic sulfate-reducing bacteria.
In Balows A, Truper HG, Dworkin M, Harder W, Schleifer KH, eds, The
Pokaryotes. A Handbook on the Biology of Bacteria: Ecophysiology, Isolation,
Identification, Applications, 2nd ed, Vol 4. Springer-Verlag, New
York, NY, pp 3352-3378.
- King JK, Harmon SM, Fu TT, Gladden JB. 2002. Mercury removal, methylmercury
formation, and sulfate reducing bacteria profiles in wetland mesocosms.
Chemosphere 46:859-870.
- US EPA. 1998. Method 1630: Methyl Mercury in Water by Distillation,
Aqueous Ethylation, Purge and Trap, and Cold Vapor Atomic Fluorescence
Spectrometry. August 1998. Washington, DC.
- Horvat M., Bloom NS, Liang L. 1993. Comparison of distillation with
other current isolation methods for the determination of methylmercury
compounds in low level environmental samples. Part I. Sediments. Anal
Chim Acta 281:135-152.
- Liang L, Horvat M, Bloom NS. 1994. An improved speciation method for
mercury by GC/CVAFS after aqueous phase ethylation and room temperature
precollection. Talanta 41:371-379.
- Bloom NS. 1989. Determination of picogram levels of methylmercury
by aqueous phase ethylation, followed by cryogenic gas chromatography
with cold vapor atomic fluorescence detection. Can J Fish Aquat Sci
46:1131-1140.
- Bloom NS, Fitzgerald WF. 1988. Determination of volatile mercury species
at the picogram level by low temperature gas chromatography with cold
vapor atomic fluorescence detection. Analytica Chimica Acta 208:151-161.
- US EPA. 1999. Method 1631, Revision B: Mercury in Water by Oxidation,
Purge and Trap, and Cold Vapor Atomic Fluorescence Spectrometry. May
1999. Washington, DC.
- Gill GA, Fitzgerald WF 1985. Mercury sampling of open ocean waters
at the picomolar level. Deep Sea Research. 32(3):287-297.
- Smith RG. 1993. Determination of mercury in environmental samples
by isotope dilution/ICPMS. Anal Chem 65:2485-2489.
- Brouwer H, Murphy TP. 1994. Diffusion Method for the Determination
of Acid-Volatile Sulfides (AVS) in Sediment. Environ Toxicol Chem
13:1273-1275.
- APHA. 1985. Standard Methods for the Examination of Water and Wastewater,
16th ed. Part 900- Microbiological Examination of Water. American Public
Health Association. Washington, DC.
- King JK, Kostka JE, Frischer ME, Saunders FM, Jahnke RA. 2001. A quantitative
relationship that demonstrates mercury methylation rates in marine sediments
are based on the community composition and activity of sulfate-reducing
bacteria Environ Sci Technol 35:2491-2496.
- Benoit JM, Gilmour CC, Mason RP. 2001b. The influence of sulfide on
solid-phase mercury bioavailability for methylation by pure cultures
of Desulfobulbus propionicus (1pr3). Environ Sci Technol
35(1):127-132.
- Oremland RS, Culbertson CW, Winfrey MR. 1991. Methylmercury
decomposition in sediments and bacterial cultures: involvement of methanogens
and sulfate reducers in oxidative demethylation. Appl Environ Microbiol
57:130137.
- Pak KR, Bartha R. 1998. Mercury methylation and demethylation in anoxic lake sediments and by strictly anaerobic bacteria. Appl Environ Microbiol 64(3):1013-1017.
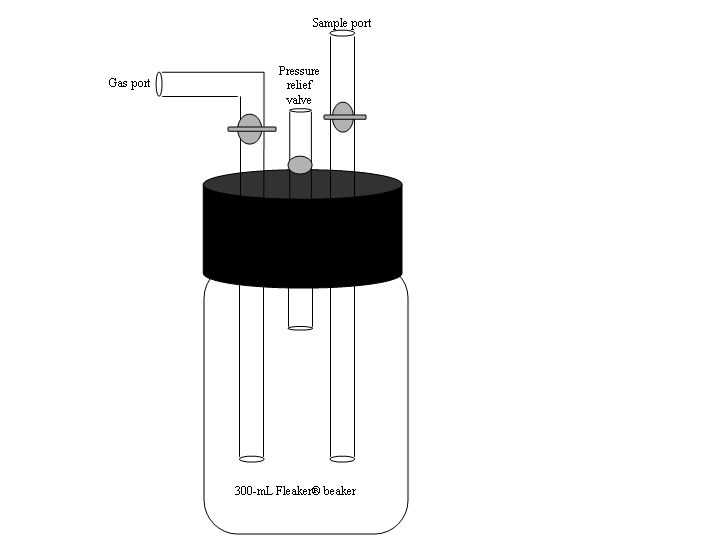
Figure 1. Sediment Slurry Reactor Vessels.
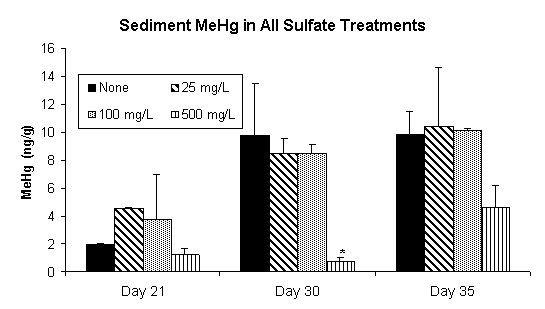
Figure 2. Sediment Methylmercury Concentrations in Sediment
Slurry Reactors. Data has been Normalized for Dry Weight.
Methylmercury is Significantly Lower (p<0.05) in the 500 mg/L
Treatment on Day 30. Data Represents the Mean of Duplicate Values.
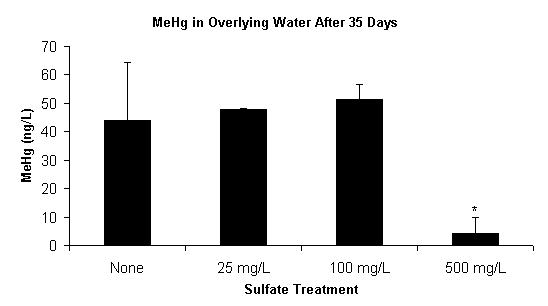
Figure 3. Dissolved Methylmercury (< 45 µm) in the Overlying
Water of the Sediment Slurry Reactors after 35 Days. Asterisk (*)Indicates
that Methylmercury Concentrations are Significantly Lower (p<0.05)
in the
500 mg/L Sulfate Treatment. Data Represents the Mean of Duplicate Values.
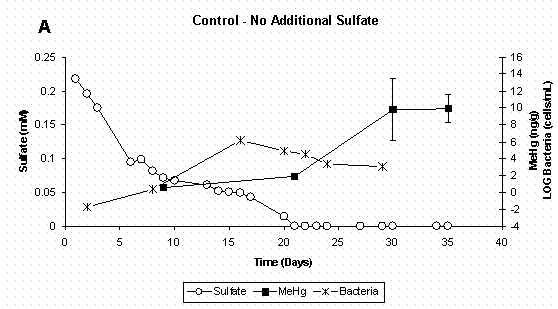
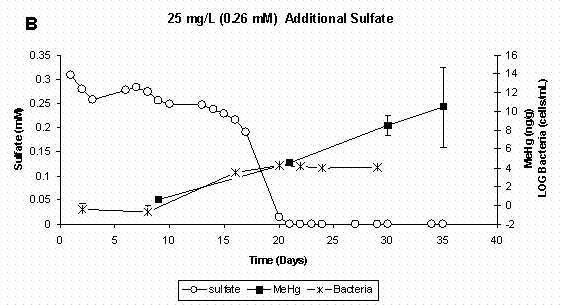
Figure 4 A&B. Dissolved Sulfate, Sediment Methylmercury, and
Bacterial Populations in Sediment Slurry Reactors. Error Bars for
Methylmercury Represent One Standard Deviation for Duplicate Samples.
Error Bars for Bacteria Represent 95% Confidence Intervals.
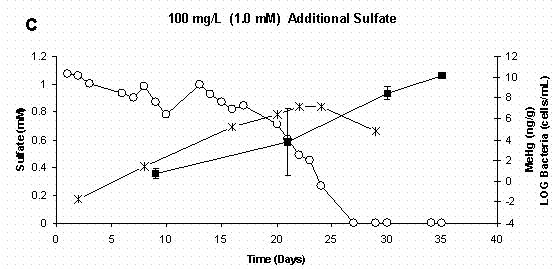
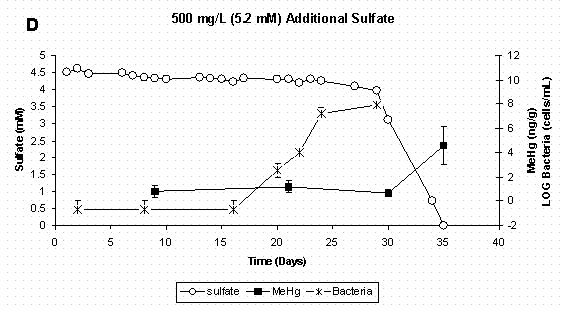
Figure 4 C&D. Dissolved Sulfate, Sediment Methylmercury, and
Bacterial Populations in Sediment Slurry Reactors. Error Bars for
Methylmercury Represent One Standard Deviation for Duplicate Samples.
Error Bars for Bacteria Represent 95% Confidence Intervals.
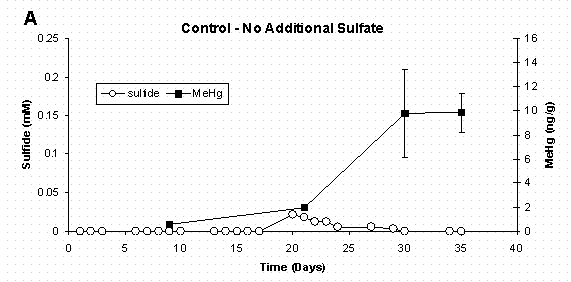
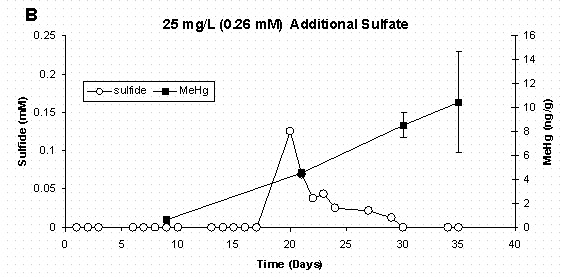
Figure 5 A&B Dissolved Sulfide and Sediment Methylmercury
Concentrations Error Bars for Methylmercury Represent
One Standard Deviation for Duplicate Samples.
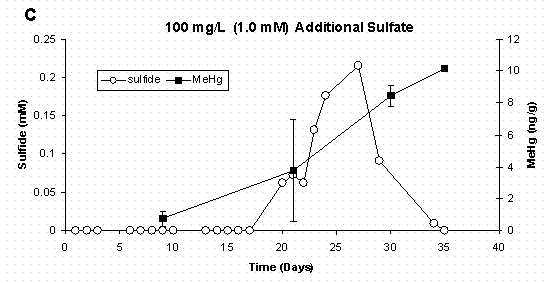
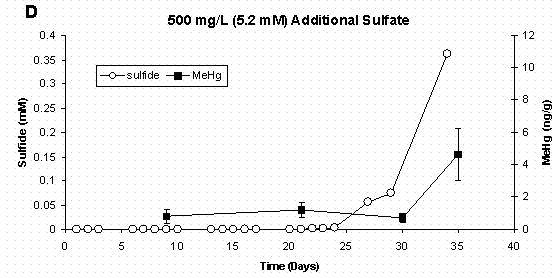
Figure 5 C&D Dissolved Sulfide and Sediment Methylmercury
Concentrations Error Bars for Methylmercury Represent
One Standard Deviation for Duplicate Samples.